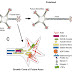
Information flows along a neuron in one direction i.e. synapse to cell body, then to axon initial segment (AIS), from their its passed to Axon and then to terminal region. Actually, neurons will receive both excitatory and inhibitory synaptic inputs on their cell bodies and dendrites. The fire of the repeated synaptic inputs will be summated and bursted out as action potential which will be taken cared by AIS. Axonal initial segment (AIS) is made up of high densities of voltage gated Na+ and K+ channels which initiate and modulate action potential. In the myelinated axons of vertebrates, the propagation of action potential is rapid along the axon through the activated cluster of sodium channels at the node of Ranvier. The action potential reaches the axon terminal and helps in release of neurotransmitters to propagate the signal across the synaptic cleft to another cell.

Fig 1: Show the sequence of information follow through the neuron from the synapses. The AIS can be seen soon after the beginning of the axon (green) which consists of node of Ranvier. Node of Ranvier is found at different regions of the axon which consists of cluster of sodium channels.
The axonal initial segment is enriched with sodium gated channels which facilitates a high sodium current density and a low action potential threshold.
The assembly of AIS is actually an intrinsic property of neuron where no extracellular or glial dependent cues are required. But the formation and localization of node of Ranvier is regulated by glial derived signals.
Ankyrin G
A cytoskeleton scaffold protein which is a master organizer of membrane domains and subcellular polarity in many cell types. This protein is restricted to AIS and AIS targeting motif located in the cytoplasmic loop of the neuron which connects the domain II and III of sodium channels binding to AnkG. Sodium channel and AnkG interaction is facilitated by Phosphorylation of AIS targeting motif by casein kinase II (CK2), which is enriched at the AIS and node of Ranvier.
Removal or silencing of AnkG protein will lead blocking the clustering of sodium channels at AIS. And also it will lead to blocking the subcellular polarization of potassium channels KCNQ2 and KCNQ3, the cell adhesion molecules neurofascin and neuronal cell adhesion molecules NrCAM, the AIS extracellular matrix and the cytoskeleton protein IV spectrin.
From this we can learn that AnkG functions as a scaffold to which all other AIS proteins are tethered directly or indirectly and consequently AnkG establishes the subcellular polarity of these molecules.
The voltage gated sodium channel Nav1.6 and Nav 1.2 are found in the distal and proximal AIS. The Kv1 family is also found primarily in the distal AIS.

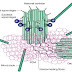
Fig 2: shows the structure of AIS
- Ion channels (Nav1.x, KCNQ2–KCNQ3 and Kv1.x)
- Neuronal cell adhesion molecule (NrCAM)
- Neurofascin 186 (NF186)
- Disintegrin and metalloproteinase domain-containing protein 22 (ADAM22)
- Transient axonal glycoprotein 1 (TAG1, also known as contactin 2)
- CASPR 2
- Extracellular matrix molecules (brevican and versican)
- Cytoskeletal scaffolds (AnkG, "IV spectrin and postsynaptic density protein 93 (PSD93))
Protein with unknown role in AIS
- Casein kinase II (CK2)
- Phosphorylated nuclear factor-κB (pNFκB)
- Phosphorylated inhibitor of κBα (pIκBα)