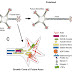
Polarized cells such as neurons have two distinct domains i.e. molecular and functional domains which have been emerged from the cell body. In neurons, axon and dendrites are two distinct functional units emerged from the cell body by the process called neuronal polarity. Axons are thin, single and long structure which will transmit the signals to the target or synaptic partner. Whereas dendrites are shorter, multiple structures which has the ability to receive signals from different regions of cells or axons.
Before getting with role of microtubules in axon and dendrite formation, we need to look into the neuronal polarity part which is already been explained in one of my post. I.e. a hippocampal neuron polarize in the presence of insulin like growth factor (IGF-I) without any cell- cell interaction or extracellular matrix. Now these polarized cells exhibit the molecular features that distinguish axon and dendrites.
So in neuronal polarity, these are the list of protein involved:
1) Kinases
2) Phosphatase
3) Small GTPase
4) Scaffolding proteins
Now what microtubule has to do with neuronal polarity?
Actually if polarity is lost, then it could be due to some modification in microtubule organization and dynamics. Hence microtubule has a very important role in neuronal polarity.
When do we say that neuronal polarization takes place?
It happens to place when there is local microtubule assembly and stabilization in one of the neurite. Change in the dynamics of the microtubule will lead to alteration in axon and dendrite specification.

Hollow cylindrical, covalent cytoskeleton polymers (24 nm diameter)
Made of tubulin proteins (α tubulin and β tubulin)
For polymerization Tubulin dimers bound to GTP
For depolymerization Tubulin dimers bound to GDP
α tubulin and β tubulin forms dimer
γ tubulin are attached at the minus end by nucleation. They form a ring at the minus which act as a cap to prevent further addition of any tubulin at the site.
Microtubule structure
Two ends of microtubules: plus end (+) and minus end (-)
At Plus end (+), dimers with GTP are assembled and GTP bound to β tubulin gets hydrolyzed to GDP through inter dimer contacts. Growth and shrinkage of microtubule takes at this end.
At minus end (-), dimers gets dissociated and polymerization at this end is prevented by cap formed by γ-tubulin ring complex. Basically, minus ends are anchored to the centrosome or may be found as free ends in some cells.
Function: cell motility, mitosis, intracellular transport, secretion, maintenance of cell shape and cell polarization.
 |
| Cycle of microtubule growth and shrinkage |
Microtubule has the ability to undergo the cycle of growth and disassembly (as shown in the fig). They never reach the steady state length, but they exist in either polymerization or depolymerization state.
There are two things happen in this cycle. One is catastrophe where depolymerization of the tubulin dimers takes. And other is rescue where the dimers polymerize together for the growth.
These two things depend on the binding of GTP at the E site (nucleotide exchangeable site) of the β tubulin. During polymerization, the E site of the β tubulin has to be bound with GTP so that α tubulin can come and bind to it. Polymerization dynamics of microtubules are important for their biological functions because they allow themselves to rapidly reorganize, differentiate spatially, temporally in accordance with cell context, and to generate pushing and pulling forces during polymerization and depolymerization.
Microtubule Associated proteins (MAPs)
These are the protein which joins the two microtubules together and form a bundle. There are two MAPs which are present in the axon and dendrites:
a) Tau protein
b) MAP2
a) Tau protein
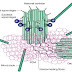
Microtubules in axon has uniform orientation with their plus ends facing the axon tip and it’s bundled up together by the protein called tau protein. It’s also present in the somato-dendritic compartment and axon region. It’s not having any essential role in axon growth and microtubule stability. This statement was actually studied in tau knock out mice where its neuron showed no significant difference with that of the wild type mice. Except in one case where the tau deficient mice showed reduction in packing density and number of microtubules in cerebellar parallel fibers, a small type of caliber axon.
But this protein is important in the case of neurite outgrowth as well as growth cone motility. Studies have been taken place in chick sensory neuron (dorsal root ganglion) where inactivation of tau protein leads to reduction in the number and length of neurite. In the growth cone, inactivation of tau protein lead to 20% decrease in the lamellopodial size and there lamellopodial motility is affected.
Tau protein plays a vital role in development of Alzheimer’s disease.
b) MAP2
Microtubules in dendrites have multiple orientations with their plus ends facing either the cell body or the dendritic tips and the bundling protein here is the MAP2. It has 3 or 4 microtubule binding domains at the COOH- terminal, and is involved in microtubule assembly and stabilization in dendrites. MAP2 is an anchoring protein of PKA in dendrites, whose loss leads to reduced amount of dendritic and total PKA and reduced activation of CREB.


Fig 3: Developed Neuron: Microtubules in Axon and dendrites are shown by magnifying the parts. Tau in axonal microtubule and MAP2 in somatic dendritic compartment. organelles such golgi apparatus, polyribosomes and Endoplasmic reticulum are localized in the dendritic region.(Nature Neuroscience review)