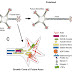
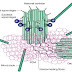
Astrocytes are star shaped glial cells which are found associated
with the neurons of the CNS and spinal cord.
- They modulate the aspects related to neural network functions.
- Express the same sets of ion channels as neurons, but in varying proportion.
- Maintenance of K+ and water homeostasis in the CNS via coordinated action of Kirl.4 and aquaporin 4 channels.
- Gap junctions form intercellular networks and help in sequestering K+ and glutamate.
- Modulation of synaptic transmission.

1. Lamellar
fibrous astrocytes
- Found in white matter
- Oriented parallel to the neuronal axons
- Higher levels of GFAP
- End feet envelops the node of Ranvier
- Found in cerebellum equipped with a large repertoire of receptors which helps in sensing the activity of synapses.
- High densities of glutamate transporters.
- Cell body spans both gray and white matter.
- Found in gray matter of CNS.
- At least one of their process is in contact with the blood vessels.
- Low input resistance.
- Very negative membrane potential.
- Prominent glutamate uptake.
- Voltage and time independent K currents.
- End feet envelops the synapses.
GFAP – Glial fibrillary acidic protein: biomarker of
Astrocytes.
Plasma membrane ion channels
1. Cx43 channels
- It’s selective for second messengers (cAMP, IP3 and calcium), amino acids (Glutamate, aspartate and taurine), Nucleotide (ADP, ATP, CTP and NAD), Energy metabolites (glucose, glucose 6 phosphate and lactate), small peptides (glutathione) and RNA (24 mer).
- Not permeable for large molecules such as nucleic acids, proteins and lipids.
2. Cx30
channels
S.No
|
Receptors
|
Transporters
|
1
|
Purine
receptors
|
Glutamate
transporter
|
2
|
mGlu
receptors
|
Glucose
transporter
|
Uptake of glutamate by glial specific
transporters:
EAAT1 (Excitatory amino acid transporters) (in rodent it’s
known as GLAST (glutamate aspartate transporter) and GLT1 (glutamate
transporter 1)) and EAAT2.
Function of Astrocytes:
(1) Astrocyte functions include modulation of synaptic
function via glutamate transporters, which convey glutamate from the synaptic
cleft into the cell.
(2) Communication between astrocytes occurs via ATP
release and binding to purine receptors on adjacent astrocytes. ATP binding
results in phospholipase C activation, with subsequent downstream activation of
inositol trisphosphate, resulting in calcium mobilization.
(3) Gap junctions contribute to an astrocyte syncytium for
the exchange of small molecules and cell–cell communication.
(4) Metabolic functions include
- The replenishment of neuronal glutamate via the glutamate glutamine cycle. (4)
- The transport of glucose from the vasculature. (5)
(6) The regulation
of blood flow is modulated by astrocyte end-feet apposing blood vessels, with
vasodilation being mediated through release of vasoactive substances.
(7) Glutamate release might occur following elevations in
intracellular calcium and the activation of other factors related to
prostaglandins.
(8) Glutamate release through hemichannels can be induced in
vitro through lowering of extracellular calcium.
(9) Glutamate binding to metabotropic glutamate receptors
activates intracellular calcium, leading to the release of vasodilatory
substances. Abbreviations: Gln, glutamine; Glu, glutamate; IP3, inositol
trisphosphate; PLC, phospholipase C.
Extra points:
Glutamate
released from neurons increases glucose trafficking in Astroglial networks by
activating AMPA (a-amino-3-hydroxy-5-methyl-4-Isoxazole propionic acid)
receptors.
Homotypic
Cx30 and Cx43 channels and heterotypic Cx43 – Cx30 channels are voltage
dependent.
Astroglial
networks supply glucose and lactate to sustain hippocampal synaptic
transmission and studies investigating the role of IP3 in hippocampal function
suggested that this molecule can induce glutamate release from gap junction
coupled astrocytes, triggering transient depolarization and epileptiform
discharges in CA1 Pyramidal neurons.
Function related to neural formation:
Control of synapse formation and
function
Regulating
number, size and shape of dendritic spines in Hippocampal CA1 pyramidal neurons
via EphA3-A4 receptors.
Adult neurogenesis
Instructing
neuronal fate commitment and by promoting proliferation of adult neural stem
cells. Moreover the effects of astrocytes are regionally specified: astrocytes
from adult hippocampus retain the potential to promote neurogenesis, but
astrocytes from adult spinal cord do not.
Astrocyte
excitation happens in two main forms:
Generated by chemical signals in neuronal
circuits(Neuron dependent excitation).
Occurs independently of neuronal input
(spontaneous excitation).
Propagation of
glutamatergic transmission via the astrocyte network.
Propagation of glutamate transmission
through the astrocyte syncytium
occurs through two prominent calcium-mediated mechanisms:
- Gap junctions
- Paracrine release of ATP
Activation
of metabotropic glutamate receptors on astrocytes following neuronal release of
glutamate results in the activation of an inositol trisphosphate (IP3) pathway,
which induces calcium release from intracellular stores. This calcium can then
be transferred to the adjacent astrocyte through connexin 43 (Cx43) gap junctions, thereby producing a calcium wave through
an astrocyte syncytium. IP3 also activates ATP release through Cx43 hemichannels. This ATP release
acts in a paracrine fashion, activating purine receptors on adjacent
astrocytes. This activation results in IP3 production, more ATP release and
intracellular calcium mobilization through a feed-forward mechanism.
Astrocytes association with Neurodegenerative Disease